Evidence for Theism
13
The Argument from Biological Teleology
Teleological arguments have explanatory force when two conditions are met: Our observations are congruent with the telos or intention of an agent and they are prohibitively improbable on the hypothesis that no such agent exists. Observing many houseflies on a wall hit by darts, for example, is congruent with the intention of a marksman to kill flies but prohibitively improbable on the hypothesis that the darts are hitting the wall at random. Therefore, the existence of a marksman is implicated. In the previous chapter, we applied teleological reasoning to the fine tuning of the physical constants and initial conditions at the Big Bang. Since this is congruent with the intention of an intelligent agent to create life, but prohibitively improbable on the hypothesis that it occurred by chance, an intelligent agent was implicated.
However, it is important to note that a fine tuned universe is a necessary but not a sufficient condition for life. [1] It merely provides a suitable substrate for life without explaining its origin and development. And this means that the telos implicated by fine tuning is not fulfilled until life appears and that the appearance of life is a second and separate explanandum. In the following paragraphs, it will be my concern to show that the riddle of improbability presented by the cosmos at large in the case of fine tuning is repeated at the molecular level in the case of abiogenesis—a fact which both completes and adds significant force to the conclusion of the previous argument since we will now be faced with one prohibitive probability balanced upon another.
Biological Explananda
Fine tuning, as already noted, does not of itself explain the enigmatic saltus from dead matter to teleological intelligence. John Locke was especially struck by this problem. “It is as impossible to conceive that ever incogitative Matter should produce a thinking intelligent Being,” he wrote, “as that nothing should of itself produce Matter.” Locke registered his puzzlement almost two hundred years before the publication of On the Origin of the Species. However, Darwin would have been of little help. Since his theory of evolution purported to explain how life developed and speciated from “one or a few simple forms,” the origin of those forms lay, and still lies, outside its explanatory boundary. [2] I will now be arguing that life instantiates three properties whose origination is as improbable on naturalism as fine tuning; namely, reproductivity, semantic information and teleology.
The Origin of Genetic Reproductivity Evolution involves the winnowing of random genetic mutations by natural selection—a process that may govern the development of organisms on Earth but which cannot explain the origin of those organisms any more than a computer program can explain the origin of electrical circuitry. Another way of expressing this point is to say that evolution presupposes the existence of self-replicating genetic material. The specific problem to be solved, therefore, is the emergence of the first genetic replicators from prebiotic matter—a problem whose difficulty is proportional to the organisation and complexity of those replicators, and that organisation and complexity is staggering indeed. [3]
The simplest conceivable biological cell capable of replication is a miniaturised factory of extraordinary complexity. A key player in its maintenance is the protein: an aggregate of amino acids which meshes with other molecules to build key structures that together regulate the metabolism of the cell, pass materials back and forth across its membranes, destroy waste and perform many other tasks. All this swarming activity is coordinated by the spiralling ladder of DNA whose sugar-and-phosphate backbone encodes instructions for the assembly of proteins from amino acids and also performs the key role in genetic replication: Its ladder is “unzipped,” copied, transcribed to RNA, and then conveyed to the amino acids which are assembled into new proteins. Two points here are critical and problematic. The first is that the errors which occur during replication produce the mutations on which natural selection operates. The second is that DNA replication cannot occur without the proteins which DNA itself constructs. It follows that evolution depends on a duality of protein manufacture and composition within the cell which poses a riddle of causality evolution cannot solve. As Robert Sharpiro explains,
|
Which came first, the chicken or the egg? DNA holds the recipe for protein construction. Yet that information cannot be retrieved or copied without the assistance of proteins. Which large molecule appeared first? Proteins—the chicken; or DNA—the egg?
|
It is in light of this paradox that Theodosius Dobzhansky has declared, "prebiological natural selection is a contradiction in terms." Whatever theory is advanced for the appearance of the first living cell, it cannot call to its aid the explanatory power of evolution.
Semantic Information Understanding the second property of living matter which naturalism cannot account for requires a familiarity with two concepts used in information theory which I will now briefly describe.
Information theory equates the amount of information transmitted by an event with the amount of uncertainty eliminated. Thus rolling a die conveys more information than flipping a coin because the former eliminates five out of six possible outcomes and the latter only one out of two. On this view a random sequence of letters and a meaningful phrase both convey information and must be individuated into the syntactic and semantic. Consider the following,
Semantic Information Understanding the second property of living matter which naturalism cannot account for requires a familiarity with two concepts used in information theory which I will now briefly describe.
Information theory equates the amount of information transmitted by an event with the amount of uncertainty eliminated. Thus rolling a die conveys more information than flipping a coin because the former eliminates five out of six possible outcomes and the latter only one out of two. On this view a random sequence of letters and a meaningful phrase both convey information and must be individuated into the syntactic and semantic. Consider the following,
|
dhcrm l chtdjf odjan rkkjfopbq
Torchlight red on sweaty faces |
The first sequence is merely syntactic. It conveys information insofar as it eliminates every other possible sequence of its length from a total of 26 characters but it has no meaning. The second sequence is semantic. In addition to eliminating every other possible sequence of its length from a total of 26 characters it also communicates a meaning. Interestingly, and significantly, the information coded in DNA is closer to the second sequence than the first. [4]
We can begin to understand this by first noting that while both of the above sequences are highly improbable only the second exhibits specification: It is specified to one of a proportionally infinitesimal number of intelligible sequences within the total set of possible permutations of letters. [5] And because this smaller set distinguishes functional from nonfunctional English sequences, and because the functionality of alphabetic sequences depends on the preexisting conventions of the English language, the smaller set qualifies as a “conditionally independent” pattern.
All of this can be tidily transposed to biological organisms. To maintain viability, the cell must manufacture specific molecular constituents whose three-dimensional shapes require equally specific arrangements of nucleotide bases. Like the letters of an alphabet, the chemical properties of DNA allow a vast number of possible sequences of nucleotide bases. And within that set of possible sequences, a proportionally infinitesimal number will produce functional proteins. Because this smaller set distinguishes functional from nonfunctional proteins, and because the functionality of nucleotide base sequences depends on the preexisting requirements of protein function, the smaller set again qualifies as a “conditionally independent” pattern. What this means is that any sequence that meets these requirements is not only improbable, like a sequence of English gibberish, but specified to a preexisting pattern, like a meaningful phrase: The nucleotide sequences in the coding regions of DNA exhibit both syntactic and semantic information. Paul Davies concludes, “The problem of how meaningful or semantic information can emerge spontaneously from a collection of mindless molecules subject to blind and purposeless forces presents a deep conceptual challenge.”
Teleology Borges, abridging Schopenhauer, said that there is something in the universe that “wants to live” and is manifest in all things. “It lies dead in minerals, dormant in plants, dreams in animals and reaches consciousness in man.” Shaw and Bergson spoke of it, respectively, as the “life force” and “élan vital.” Schopenhauer himself called it wille. “Here,” concluded Borges, “we have the explanation of that line of Aquinas: Intellectus naturaliter desiderat esse semper: ‘The mind naturally desires to exist forever.’”
The felicity of Borges’ phrasing masks an infelicity of philosophy. If a property is “dead” in some object can that object be said to instantiate it? In his discussion of the subject, Antony Flew draws our attention to the obvious point. “Living matter,” he writes, “possesses a goal-directed organisation that is nowhere present in the matter that preceded it.” Scientists from Haldane to Ayala concede that teleological language is ineliminable in evolutionary biology—at the same time as they bridle at its affinity to natural theology. “Teleology is like a mistress to a biologist,” said Haldane. “He cannot live without her but he's unwilling to be seen with her in public."
The third explanandum, then, is how the prebiotic soup of the early Earth gave rise to matter so radically different from itself; matter that resists description in nonteleological terms; that possesses intrinsic ends, goals, purposes—pursues a good of its own. Flew thinks this question poses a deep philosophical challenge to biology that has been largely overlooked. "Most studies on the origin of life," he explains, "are carried out by scientists who rarely attend to the philosophical dimension of their findings. Philosophers, on the other hand, have said little on the nature and origin of life."
Explanatory Options
We have seen that the leap from chemistry to biology resists naturalistic explanation on account of three properties: The ability of living matter to replicate, convey and process semantic information and pursue its own ends. In what follows I will briefly discuss the inviability of the only explanatory options available to the naturalist (chance, prebiotic evolution and chemical necessity) before defending the teleological explanation.
Chance Almost all serious origin-of-life researchers now consider sheer chance an inadequate causal explanation for the origin of life. To understand why, consider the probability of even one short protein molecule 100 amino acids in length forming at random under ideal prebiotic conditions.
Amino acids must form a chemical bond known as a peptide bond in the protein chain. Many other types of chemical bonds are possible; in fact, peptide and nonpeptide bonds occur with equal probability. The probability of a chain of 100 amino acids in which all bonds are peptide bonds forming at random is about 1:10^30. Moreover, every amino acid found in proteins, with one exception, has two distinct mirror images of itself called optical isomers. Functioning proteins tolerate only left-handed amino acids but the right-handed and left-handed isomers occur in chemical reactions with equal frequency. The probability of a peptide chain of 100 left-handed amino acids forming at random is about 1:10^30. Putting this together: The probability of a 100-amino-acid-length chain in which all bonds are peptide bonds and all amino acids are left-handed is about 1:10^60. Functional proteins, finally, must link up in a specific sequential arrangement. Changing even one amino acid results in loss of function. [6] It has been calculated that the probability of attaining a particular protein 100 amino acids long is about 1:10^130—overwhelmingly and prohibitively improbable through the operation of chance alone on the chemical soup of the early earth.
And this, recall, is for a single short protein under hypothetically optimal prebiotic conditions. Calculations for the chance assembly of an organism with the minimal complexity to sustain and reproduce itself under more realistic conditions suggest odds of about 1:10^150—an event far exceeding even the probabilistic resources of a concentrated solution of amino acids the size of the observable universe.
Prebiotic Natural Selection Dawkins and Kuppers, conceding all the above, have used computer models in an attempt to demonstrate how “prebiotic natural selection” might bring the probabilities to within feasible limits. Each chooses a target sequence to represent a desired functional polymer. After creating a crop of randomly constructed sequences and generating variations among them at random, the computer program selects those sequences that match the target sequence most closely. In this way the target phrase is soon reached. The problem is that, unlike a human agent designing a computer program, molecules in situ do not have a target phrase “in mind.”
Discussions of this problem have routinely appealed to the analogy of n number of monkeys hitting typewriter keys at random over varying periods of time—one often used to illustrate the emergence of complexity from random processes. In its most familiar formulation, a monkey with infinite time and paper produces the complete works of Shakespeare. But the idea, while it seems apposite to the task of explaining the origin of life, actually serves to demonstrate the impossibility of even modest complexity emerging from randomness as soon as any sort of time constraint is imposed. And this is a problem because time constraints obviously apply.
John Lennox, an Oxford professor of mathematics, is comfortably at home as he walks us through the various calculations. A representative example: To type out Hamlet a monkey hitting one random key per nano second would need a length of time next to which the estimated life of the universe would appear insignificant by comparison. And as Hoyle and Wickramasinghe inform us, we cannot bring the probability to within feasible limits by multiplying the monkeys because the observable universe is simply not large enough to contain the number of monkeys required.
Applying the analogy to Dawkins and Kuppers, they have circumvented the above problem by teaching their monkeys a rule: For all monkeys there is a target phrase and for each monkey a target letter. As paraphrased by Lennox,
We can begin to understand this by first noting that while both of the above sequences are highly improbable only the second exhibits specification: It is specified to one of a proportionally infinitesimal number of intelligible sequences within the total set of possible permutations of letters. [5] And because this smaller set distinguishes functional from nonfunctional English sequences, and because the functionality of alphabetic sequences depends on the preexisting conventions of the English language, the smaller set qualifies as a “conditionally independent” pattern.
All of this can be tidily transposed to biological organisms. To maintain viability, the cell must manufacture specific molecular constituents whose three-dimensional shapes require equally specific arrangements of nucleotide bases. Like the letters of an alphabet, the chemical properties of DNA allow a vast number of possible sequences of nucleotide bases. And within that set of possible sequences, a proportionally infinitesimal number will produce functional proteins. Because this smaller set distinguishes functional from nonfunctional proteins, and because the functionality of nucleotide base sequences depends on the preexisting requirements of protein function, the smaller set again qualifies as a “conditionally independent” pattern. What this means is that any sequence that meets these requirements is not only improbable, like a sequence of English gibberish, but specified to a preexisting pattern, like a meaningful phrase: The nucleotide sequences in the coding regions of DNA exhibit both syntactic and semantic information. Paul Davies concludes, “The problem of how meaningful or semantic information can emerge spontaneously from a collection of mindless molecules subject to blind and purposeless forces presents a deep conceptual challenge.”
Teleology Borges, abridging Schopenhauer, said that there is something in the universe that “wants to live” and is manifest in all things. “It lies dead in minerals, dormant in plants, dreams in animals and reaches consciousness in man.” Shaw and Bergson spoke of it, respectively, as the “life force” and “élan vital.” Schopenhauer himself called it wille. “Here,” concluded Borges, “we have the explanation of that line of Aquinas: Intellectus naturaliter desiderat esse semper: ‘The mind naturally desires to exist forever.’”
The felicity of Borges’ phrasing masks an infelicity of philosophy. If a property is “dead” in some object can that object be said to instantiate it? In his discussion of the subject, Antony Flew draws our attention to the obvious point. “Living matter,” he writes, “possesses a goal-directed organisation that is nowhere present in the matter that preceded it.” Scientists from Haldane to Ayala concede that teleological language is ineliminable in evolutionary biology—at the same time as they bridle at its affinity to natural theology. “Teleology is like a mistress to a biologist,” said Haldane. “He cannot live without her but he's unwilling to be seen with her in public."
The third explanandum, then, is how the prebiotic soup of the early Earth gave rise to matter so radically different from itself; matter that resists description in nonteleological terms; that possesses intrinsic ends, goals, purposes—pursues a good of its own. Flew thinks this question poses a deep philosophical challenge to biology that has been largely overlooked. "Most studies on the origin of life," he explains, "are carried out by scientists who rarely attend to the philosophical dimension of their findings. Philosophers, on the other hand, have said little on the nature and origin of life."
Explanatory Options
We have seen that the leap from chemistry to biology resists naturalistic explanation on account of three properties: The ability of living matter to replicate, convey and process semantic information and pursue its own ends. In what follows I will briefly discuss the inviability of the only explanatory options available to the naturalist (chance, prebiotic evolution and chemical necessity) before defending the teleological explanation.
Chance Almost all serious origin-of-life researchers now consider sheer chance an inadequate causal explanation for the origin of life. To understand why, consider the probability of even one short protein molecule 100 amino acids in length forming at random under ideal prebiotic conditions.
Amino acids must form a chemical bond known as a peptide bond in the protein chain. Many other types of chemical bonds are possible; in fact, peptide and nonpeptide bonds occur with equal probability. The probability of a chain of 100 amino acids in which all bonds are peptide bonds forming at random is about 1:10^30. Moreover, every amino acid found in proteins, with one exception, has two distinct mirror images of itself called optical isomers. Functioning proteins tolerate only left-handed amino acids but the right-handed and left-handed isomers occur in chemical reactions with equal frequency. The probability of a peptide chain of 100 left-handed amino acids forming at random is about 1:10^30. Putting this together: The probability of a 100-amino-acid-length chain in which all bonds are peptide bonds and all amino acids are left-handed is about 1:10^60. Functional proteins, finally, must link up in a specific sequential arrangement. Changing even one amino acid results in loss of function. [6] It has been calculated that the probability of attaining a particular protein 100 amino acids long is about 1:10^130—overwhelmingly and prohibitively improbable through the operation of chance alone on the chemical soup of the early earth.
And this, recall, is for a single short protein under hypothetically optimal prebiotic conditions. Calculations for the chance assembly of an organism with the minimal complexity to sustain and reproduce itself under more realistic conditions suggest odds of about 1:10^150—an event far exceeding even the probabilistic resources of a concentrated solution of amino acids the size of the observable universe.
Prebiotic Natural Selection Dawkins and Kuppers, conceding all the above, have used computer models in an attempt to demonstrate how “prebiotic natural selection” might bring the probabilities to within feasible limits. Each chooses a target sequence to represent a desired functional polymer. After creating a crop of randomly constructed sequences and generating variations among them at random, the computer program selects those sequences that match the target sequence most closely. In this way the target phrase is soon reached. The problem is that, unlike a human agent designing a computer program, molecules in situ do not have a target phrase “in mind.”
Discussions of this problem have routinely appealed to the analogy of n number of monkeys hitting typewriter keys at random over varying periods of time—one often used to illustrate the emergence of complexity from random processes. In its most familiar formulation, a monkey with infinite time and paper produces the complete works of Shakespeare. But the idea, while it seems apposite to the task of explaining the origin of life, actually serves to demonstrate the impossibility of even modest complexity emerging from randomness as soon as any sort of time constraint is imposed. And this is a problem because time constraints obviously apply.
John Lennox, an Oxford professor of mathematics, is comfortably at home as he walks us through the various calculations. A representative example: To type out Hamlet a monkey hitting one random key per nano second would need a length of time next to which the estimated life of the universe would appear insignificant by comparison. And as Hoyle and Wickramasinghe inform us, we cannot bring the probability to within feasible limits by multiplying the monkeys because the observable universe is simply not large enough to contain the number of monkeys required.
Applying the analogy to Dawkins and Kuppers, they have circumvented the above problem by teaching their monkeys a rule: For all monkeys there is a target phrase and for each monkey a target letter. As paraphrased by Lennox,
|
Each time a monkey hits a letter, the letter it types is compared with its target letter—a highly non-random process. This comparison, of course, has to be done by some mechanism such as a computer—or a Head Monkey, as mathematician David Berlinski delightfully suggests. If the monkey has typed its target letter the comparison mechanism retains that letter and the monkey stops typing—another highly non-random process. If not, the monkey is allowed to go on randomly typing until it gets its target letter.
|
Consider for a moment both that "target phrase" and "target letter." This is what has happened: In an attempt to tame the disqualifying improbabilities involved in genetic self-assembly and so remove the suggestion of intelligent agency, Dawkins and Kuppers simply introduce an element of intelligent agency. Berlinski, in a much discussed article, describes this as "an achievement in self-deception" since, "the mechanism of deliberate design, purged by Darwinian theory on the level of the organism, has reappeared in the description of natural selection itself."
Chemical Necessity Those committed to finding a naturalistic explanation for the origin of life must turn to physical or chemical necessity. The proponent of this solution begins by noting that natural forces regularly produce order. Gravitational energy, for instance, will produce vortices in a draining bathtub; electrostatic forces will draw sodium and chloride ions together into ordered patterns within a crystal of salt. It seems reasonable enough to suppose that, in a like case, amino acids with special affinities for each other might arrange themselves to form proteins.
There are two problems to note.
The first is that there are no chemical bonds between bases along the axis of the DNA molecule and yet it is precisely along this axis that genetic information is coded. This can be seen in the diagram of bonding relationships below.
Sugars are designated by pentagons and phosphates by circled Ps. These are linked chemically—represented by solid lines. Meanwhile, nucleotide bases (As, Ts, Gs and Cs) are bonded to the sugar-phosphate backbone and linked by hydrogen bonds—the dotted lines. Note, however, an absence of vertical lines between the squares. This is because there are no chemical bonds between the nucleotide bases along the message-bearing spine of the helix. As magnetic letters on a refrigerator door can be combined and recombined in any of various ways to produce specific sequences, so nucleotide bases can attach to any site on the DNA backbone with equal facility.
Chemical Necessity Those committed to finding a naturalistic explanation for the origin of life must turn to physical or chemical necessity. The proponent of this solution begins by noting that natural forces regularly produce order. Gravitational energy, for instance, will produce vortices in a draining bathtub; electrostatic forces will draw sodium and chloride ions together into ordered patterns within a crystal of salt. It seems reasonable enough to suppose that, in a like case, amino acids with special affinities for each other might arrange themselves to form proteins.
There are two problems to note.
The first is that there are no chemical bonds between bases along the axis of the DNA molecule and yet it is precisely along this axis that genetic information is coded. This can be seen in the diagram of bonding relationships below.
Sugars are designated by pentagons and phosphates by circled Ps. These are linked chemically—represented by solid lines. Meanwhile, nucleotide bases (As, Ts, Gs and Cs) are bonded to the sugar-phosphate backbone and linked by hydrogen bonds—the dotted lines. Note, however, an absence of vertical lines between the squares. This is because there are no chemical bonds between the nucleotide bases along the message-bearing spine of the helix. As magnetic letters on a refrigerator door can be combined and recombined in any of various ways to produce specific sequences, so nucleotide bases can attach to any site on the DNA backbone with equal facility.
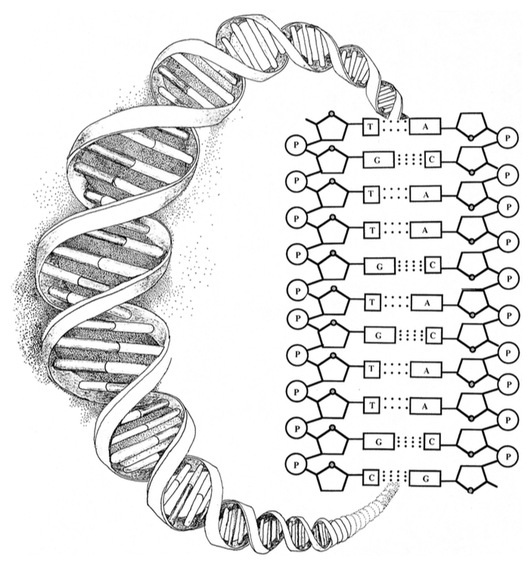
Indeed, there are no significant differential affinities between any of the four bases and the binding sites along the sugar-phosphate backbone. It follows that physical bonding affinities cannot explain the sequentially specific arrangement of nucleotide bases in DNA. Notes Carl Woese,
|
The coding, mechanistic, and evolutionary facets of the problem now became separate issues. The idea that gene expression, like gene replication, was underlain by some fundamental physical principle is gone.
|
The second problem with this explanatory option is that if chemical affinities did determine the arrangement of the bases, such affinities would dramatically diminish the capacity of DNA to carry information. Information theory, recall, equates the reduction of uncertainty with the transmission of information. The information-bearing capacity of DNA therefore requires physical and chemical contingency. Consider what would happen if each nucleotide base interacted by chemical necessity along the information-bearing axis of DNA; suppose, that is, that every time adenine occurred in a growing genetic sequence, it attracted cytosine to it; and every time guanine appeared, thymine followed. The result would be repetition and redundancy that was ordered but lacking in both information and complexity—much like the arrangement of atoms in crystals. [7]
"Whatever may be the origin of a DNA," concludes Meyer, "it can function as a code only if its order is not due to the forces of potential energy. It must be as physically indeterminate as the sequence of words is on a printed page."
Intelligent Agency Paul Davies, a physicist and origin of life researcher, thinks that the failure of science to explain the origin of life may be due to a category error; that is, he thinks that it is a problem for information theorists that has been left in the hands of chemists. “Chemistry,” he explains, “is about substances and how they react, whereas biology appeals to information and organisation.” Davies invites us to think of a computer. “Attempts at chemical synthesis focus on the hardware—the chemical substrate of life—but ignore the software—the informational aspect.” He then suggests a new perspective on the problem: The answer to the origin of life does not lie in an elusive chemical transformation but a transformation in the organisation of information flow.
In physical systems the parts determine the operation of the whole. Davies says this can be understood as, “a flow of information from the bottom up.” A meteorologist who wishes to predict the weather, for example, will begin with local conditions (temperature, air pressure, and so on) and then calculate how the system as a whole will behave. In living organisms, bottom-up information flow mingles with top-down information flow—insofar as what happens at the local level can depend on the global environment and vice versa. Citing mathematical models produced with the help of an astrobiologist, Davies concludes, “The key transition on the road to life occurred when top-down information flow first predominated. We think it may have happened suddenly, analogously to a heated gas abruptly bursting into flame.”
The image is suggestive.
Is there any known entity with the causal powers to create large amounts of specified information in this way? There is. “The creation of new information,” notes Quastler, “is habitually associated with conscious activity.” This is a precept for which experience provides amble evidence. The information on a computer screen, for example, originates in the mind of a software engineer; the information on a page, in the mind of a writer. And it holds, importantly, for other forms of specified complexity. The ultimate explanation for the meshing gears of a watch is in the intelligent agency of a horologist. Insurance-fraud investigators detect “cheating patterns” that distinguish intentional manipulation from accident; cryptographers distinguish between random signals and those that carry encoded messages. Inferring agency from specified complexity is a familiar and coherent mode of reasoning.
Evaluating the Explanatory Options
We have seen that chance is inviable on grounds of prohibitive improbability. It cannot account for the origin of a single protein—let alone a functioning molecule. Prebiotic evolution, meanwhile, is explanatorily bankrupt because it smuggles design into natural selection. And chemical necessity both fails to explain the genetic code, which is not determined by bonding, and thwarts the information carrying capacity of DNA. It is notable that all three explanatory options available to the naturalist fall at the first stile: Not one of them can account for the origination of cellular molecules. They all come to grief before semantic information and teleology even enter the discussion.
The hypothesis of intelligent agency, on the other hand, tidily explains all three properties of life and life's origin. It suggests that if there is no God it is unreasonably improbable that teleological entities capable of replicating themselves and processing semantic information will emerge from the prebiotic soup of the early Earth while if there is a God it is highly probable that this will occur. The origin of life, the argument suggests, is powerful inductive evidence for the telos of an intelligent agent—complementing, completing and adding considerable force to the previous argument from cosmic teleology. [8]
Flew, who famously renounced his atheism on review of the foregoing arguments, was moved to conclude: "Intelligence, rather than emerging as a late outgrowth of the evolution of life, has always existed as the matrix and substrate of physical reality."
Conclusion
The foregoing discussion can now be formalised into an abductive syllogism,
"Whatever may be the origin of a DNA," concludes Meyer, "it can function as a code only if its order is not due to the forces of potential energy. It must be as physically indeterminate as the sequence of words is on a printed page."
Intelligent Agency Paul Davies, a physicist and origin of life researcher, thinks that the failure of science to explain the origin of life may be due to a category error; that is, he thinks that it is a problem for information theorists that has been left in the hands of chemists. “Chemistry,” he explains, “is about substances and how they react, whereas biology appeals to information and organisation.” Davies invites us to think of a computer. “Attempts at chemical synthesis focus on the hardware—the chemical substrate of life—but ignore the software—the informational aspect.” He then suggests a new perspective on the problem: The answer to the origin of life does not lie in an elusive chemical transformation but a transformation in the organisation of information flow.
In physical systems the parts determine the operation of the whole. Davies says this can be understood as, “a flow of information from the bottom up.” A meteorologist who wishes to predict the weather, for example, will begin with local conditions (temperature, air pressure, and so on) and then calculate how the system as a whole will behave. In living organisms, bottom-up information flow mingles with top-down information flow—insofar as what happens at the local level can depend on the global environment and vice versa. Citing mathematical models produced with the help of an astrobiologist, Davies concludes, “The key transition on the road to life occurred when top-down information flow first predominated. We think it may have happened suddenly, analogously to a heated gas abruptly bursting into flame.”
The image is suggestive.
Is there any known entity with the causal powers to create large amounts of specified information in this way? There is. “The creation of new information,” notes Quastler, “is habitually associated with conscious activity.” This is a precept for which experience provides amble evidence. The information on a computer screen, for example, originates in the mind of a software engineer; the information on a page, in the mind of a writer. And it holds, importantly, for other forms of specified complexity. The ultimate explanation for the meshing gears of a watch is in the intelligent agency of a horologist. Insurance-fraud investigators detect “cheating patterns” that distinguish intentional manipulation from accident; cryptographers distinguish between random signals and those that carry encoded messages. Inferring agency from specified complexity is a familiar and coherent mode of reasoning.
Evaluating the Explanatory Options
We have seen that chance is inviable on grounds of prohibitive improbability. It cannot account for the origin of a single protein—let alone a functioning molecule. Prebiotic evolution, meanwhile, is explanatorily bankrupt because it smuggles design into natural selection. And chemical necessity both fails to explain the genetic code, which is not determined by bonding, and thwarts the information carrying capacity of DNA. It is notable that all three explanatory options available to the naturalist fall at the first stile: Not one of them can account for the origination of cellular molecules. They all come to grief before semantic information and teleology even enter the discussion.
The hypothesis of intelligent agency, on the other hand, tidily explains all three properties of life and life's origin. It suggests that if there is no God it is unreasonably improbable that teleological entities capable of replicating themselves and processing semantic information will emerge from the prebiotic soup of the early Earth while if there is a God it is highly probable that this will occur. The origin of life, the argument suggests, is powerful inductive evidence for the telos of an intelligent agent—complementing, completing and adding considerable force to the previous argument from cosmic teleology. [8]
Flew, who famously renounced his atheism on review of the foregoing arguments, was moved to conclude: "Intelligence, rather than emerging as a late outgrowth of the evolution of life, has always existed as the matrix and substrate of physical reality."
Conclusion
The foregoing discussion can now be formalised into an abductive syllogism,
|
The surprising fact p is observed
If r were the case, p would follow as a matter of course Therefore, probably, r |
The surprising fact p is the origin of life. And in discussing the completing hypotheses for r, theism has clearly emerged as an inference to the best explanation. On the basis of the four arguments so far given, we are rationally obligated to conclude that there exists an uncaused, eternal, changeless, timeless, immaterial and unimaginably powerful agent who by an act of free will brought the universe into being with the goal of creating intelligent life on Earth.
[1] A necessary condition is one that is required before some state of affairs can obtain; a sufficient condition is one that ensures that some state of affairs obtains. Thus in the case of triangles: Being a polygon is a necessary condition; being a three-sided polygon is a sufficient condition.
[2] In Darwin’s own time biologists assumed that the essence of life was a chemically simple substance called “protoplasm” and the cell an undifferentiated globule containing it. Theories of abiogenesis accordingly envisioned life arising with relative ease from a process of chemical “autogeny” that comprised only one or two steps. Such conjectures, of course, reflected a gross ignorance of the complexity of the cell.
[3] Most of the technical details that follow are taken from DNA and the Origin of Life: Information, Specification and Explanation by Stephen Meyer.
[4] The word “semantic” here carries a stronger conception of information than that of information-theorists and a slightly weaker conception that of linguists and ordinary users: While both natural languages and DNA base sequences are specified only natural language conveys subjective meaning.
[5] Just how infinitesimal readers of Borges’ The Library of Babel will appreciate.
[6] “A topoisomerase,” notes Meyer, “can no more perform the job of a polymerase than a spoon could perform the function of a corkscrew.”
[7] Imagine, by way of analogy, the difficulty you would have communicating your thoughts using a word processing program with the following feature. If you type an H, the only possible letter that may succeed it is an R; and if you type an R, the only possible letter that may succeed it is an S—and so on, for the remaining 24 letters.
[8] The inference to agency from the integrated complexity of the physical world is not a proof of the existence of God—these, as already noted, do not exist outside of mathematics and symbolic logic. But nor is it a fallacious argument from ignorance—or a “God of the Gaps.” Arguments from ignorance occur when evidence against a proposition p is offered as the sole grounds for accepting some alternative proposition r. The foregoing argument does not commit this fallacy.
[2] In Darwin’s own time biologists assumed that the essence of life was a chemically simple substance called “protoplasm” and the cell an undifferentiated globule containing it. Theories of abiogenesis accordingly envisioned life arising with relative ease from a process of chemical “autogeny” that comprised only one or two steps. Such conjectures, of course, reflected a gross ignorance of the complexity of the cell.
[3] Most of the technical details that follow are taken from DNA and the Origin of Life: Information, Specification and Explanation by Stephen Meyer.
[4] The word “semantic” here carries a stronger conception of information than that of information-theorists and a slightly weaker conception that of linguists and ordinary users: While both natural languages and DNA base sequences are specified only natural language conveys subjective meaning.
[5] Just how infinitesimal readers of Borges’ The Library of Babel will appreciate.
[6] “A topoisomerase,” notes Meyer, “can no more perform the job of a polymerase than a spoon could perform the function of a corkscrew.”
[7] Imagine, by way of analogy, the difficulty you would have communicating your thoughts using a word processing program with the following feature. If you type an H, the only possible letter that may succeed it is an R; and if you type an R, the only possible letter that may succeed it is an S—and so on, for the remaining 24 letters.
[8] The inference to agency from the integrated complexity of the physical world is not a proof of the existence of God—these, as already noted, do not exist outside of mathematics and symbolic logic. But nor is it a fallacious argument from ignorance—or a “God of the Gaps.” Arguments from ignorance occur when evidence against a proposition p is offered as the sole grounds for accepting some alternative proposition r. The foregoing argument does not commit this fallacy.
